ピーナッツアレルギー研究用ツール
IgEを介した食物アレルギーであるピーナッツに対する即時型アレルギー反応は、長年にわたり公衆衛生上の大きな懸念となっており、特に西欧諸国ではピーナッツアレルギーが成人期まで持続しています。現在のところ、ピーナッツを避けることがアレルギー患者にとっての唯一の選択肢となっています(1)。ピーナッツ中には、Ara h1、Ara h2、Ara h3、およびAra h6などの11種類の主要なアレルゲンが含まれています。特に2SアルブミンであるAra h2およびAra h6は、アレルギー反応の発症に大きく寄与します(2)。
ピーナッツアレルギーの病因の研究や、新しい治療法の評価にはマウスのピーナッツアレルギーモデルが用いられます。本モデルの評価では、ピーナッツ粗抽出物(Crude Peanut Extract;CPE)または各精製Araアレルゲンの投与によって惹起される、血清中のアレルゲンに対するIgEおよびIgG抗体などの体液性免疫応答、T細胞媒介性免疫応答に関連するサイトカインレベル、体温やアナフィラキシー症状が用いられています。疾患モデルで観察されるこれらの指標の変化は、アレルギー反応の発症に対する防御効果の評価に有用です(3-9)。
Chondrex社では、マウスピーナッツアレルギーモデルを惹起できるCPEおよび抗CPE IgEモノクローナル抗体を提供しております。さらに、CPEに対する体液性免疫応答の評価用のマウス抗CPE抗体アッセイキットや、サイトカイン・ケモカインELISAキットも提供しております。
製品ラインアップ
| 詳細 | メーカー | 製品番号 | 製品名 | 容量 | 価格 | 在庫情報 | 保存温度 | 法規制 | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chondrex | 3056 | Mouse Anti-Crude Peanut Extract (CPE) IgG Antibody Assay Kit | 1KT | ¥104,000 | 問合せ | -20℃ | ||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Chondrex | 3057 | Mouse Anti-Crude Peanut Extract (CPE) IgG1 Antibody Assay Kit | 1KT | ¥104,000 | 問合せ | -20℃ | ||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Chondrex | 3058 | Mouse Anti-Crude Peanut Extract (CPE) IgG2a Antibody Assay Kit | 1KT | ¥104,000 | 問合せ | -20℃ | ||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Chondrex | 3059 | Mouse Anti-Crude Peanut Extract (CPE) IgG2b Antibody Assay Kit | 1KT | ¥104,000 | 問合せ | -20℃ | ||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Chondrex | 3060 | Mouse Anti-Crude Peanut Extract (CPE) IgG3 Antibody Assay Kit | 1KT | ¥104,000 | 問合せ | -20℃ | ||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Chondrex | 3061 | Mouse Anti-Crude Peanut Extract (CPE) IgA Antibody Assay Kit | 1KT | ¥104,000 | 問合せ | -20℃ | ||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Chondrex | 3062 | Mouse Anti-Crude Peanut Extract (CPE) IgM Antibody Assay Kit | 1KT | ¥104,000 | 問合せ | -20℃ | ||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Chondrex | 3063 | Mouse Anti-Crude Peanut Extract (CPE) IgE Antibody Assay Kit | 1KT | ¥104,000 | 問合せ | -20℃ | ||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Chondrex | 3069 | Crude Peanut Extract | 10MG | ¥38,000 | 問合せ | -20℃ | - | |||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Chondrex | 3070 | Mouse Anti-Crude Peanut Extract (CPE) IgE Antibody 2G11G7 | 3MG | ¥153,000 | 問合せ | -20℃ | - | |||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
アレルギーモデルマウスにおけるIgE, IgE受容体および肥満細胞
食物アレルギーの免疫学的な特徴として、摂取した食物成分に対する過剰な反応である即時型過敏反応があります。このアレルギー反応では、アレルゲンに対するIgE抗体が重要な役割を担っています。IgE抗体は、高親和性のFcεRIと低親和性のFcεRIIという2種類のIgE受容体に結合することができます。FcεRIは、主に肥満細胞、好塩基球、樹状細胞に発現しています。IgE抗体およびアレルゲン(およびIgG抗体)によりFcεRIが架橋されると、受容体が活性化され、細胞内シグナル伝達が開始されます。活性化されたシグナル伝達により、細胞の脱顆粒とアミン(ヒスタミン)、プロテオグリカン、プロテアーゼ、リソソーム酵素(b-ヘキソサミニダーゼ)、脂質メディエーター、サイトカイン、およびケモカイン(GM-CSF、IL-1b、IL-8、IL-13、MCP-1)(11,12)などの放出が引き起こされます(図1a, 図1b)。
マウスモデルでは食物アレルゲン感作後の即時型過敏反応およびアナフィラキシーの発症により、IgE抗体、FcεRI、および肥満細胞の役割が検証されています(13)。ラット好塩基球性白血病細胞(RBL-2H3)は肥満細胞と同等の表現型を持っており、アレルゲン、IgE抗体、FceRI受容体、およびその後のシグナル伝達などに関連するin vitro研究に使用されています(14)。
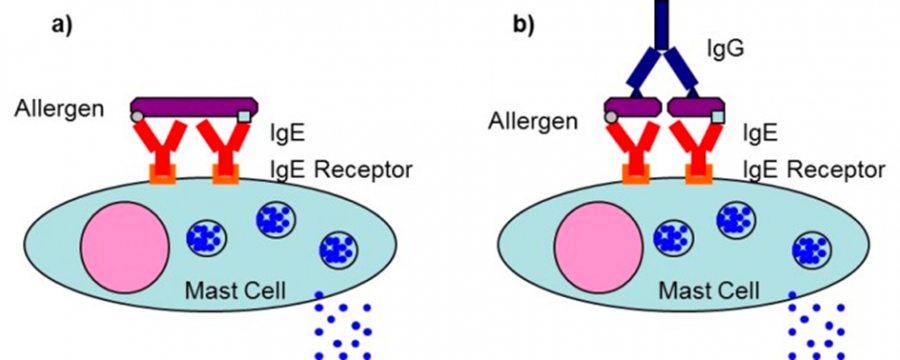
a) 単一のアレルゲンによる架橋、b) IgG抗体が結合した2つのアレルゲン分子をによる架橋
肥満細胞上のIgE受容体に結合した隣接する2つのIgE抗体が多価の単一アレルゲンを補足することにより、2つのIgE受容体が架橋され、肥満細胞の脱顆粒(活性化)が起こる(図1a)。
もしくは、IgG抗体とアレルゲンの免疫複合体も肥満細胞上のIgE抗体に補足され、IgE受容体を架橋することにより、肥満細胞を脱顆粒させることができる(図1b)。
したがって、アレルギー反応の発現には、アレルゲンに対するIgE抗体だけでなく、IgG抗体も重要な役割を担っていると考えられる。
ピーナッツアレルギーモデル
マウスピーナッツアレルギーモデルは、ピーナッツ粗抽出物(CPE)をコレラ毒素と共に2回経口投与した後に再度、経口投与でCPEを感作することで誘発し、血清中の抗CPE IgEおよびIgG抗体、T細胞性の免疫応答に関連するサイトカインレベル、体温やアナフィラキシーなどの臨床的徴候により評価が可能です(10)。
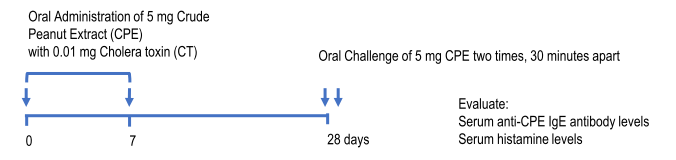
Mouse Anti-Crude Peanut Extract (CPE) IgE Antibody(Clone:2G11G7)の評価
Chondrex社ではin vitroおよびin vivo研究用の抗CPE抗体としてClone:2G11G7をご提供しております。本クローンの特性は以下の通り様々な方法で評価されております。
1) イムノブロット分析
ピーナッツ粗抽出物(CPE)のタンパク質を還元、および非還元条件下で12% SDS-PAGEにより分析しました。使用したCPEは、Ara h1、h2、h3、およびh6を含む複数のタンパク質バンドを示しました(図2)。次に、CPEに対する4つのマウスIgEモノクローナル抗体(mAb)の抗原を、CPEを用いてウエスタンブロットで分析しました。非還元条件下では6E10C12、6B7B10および5E7B12は68kDaのタンパク質を認識し、2G11G7 mAbは63kDaおよび60kDaのタンパク質を認識しました。この結果は、SDSによる変性還元条件下で100℃でCPEを変性させた後でも、それらのエピトープが安定であることを示唆しています。しかし、還元条件下では、6B7B10のみが68kDaと63kDaのタンパク質を認識しました(図2)。
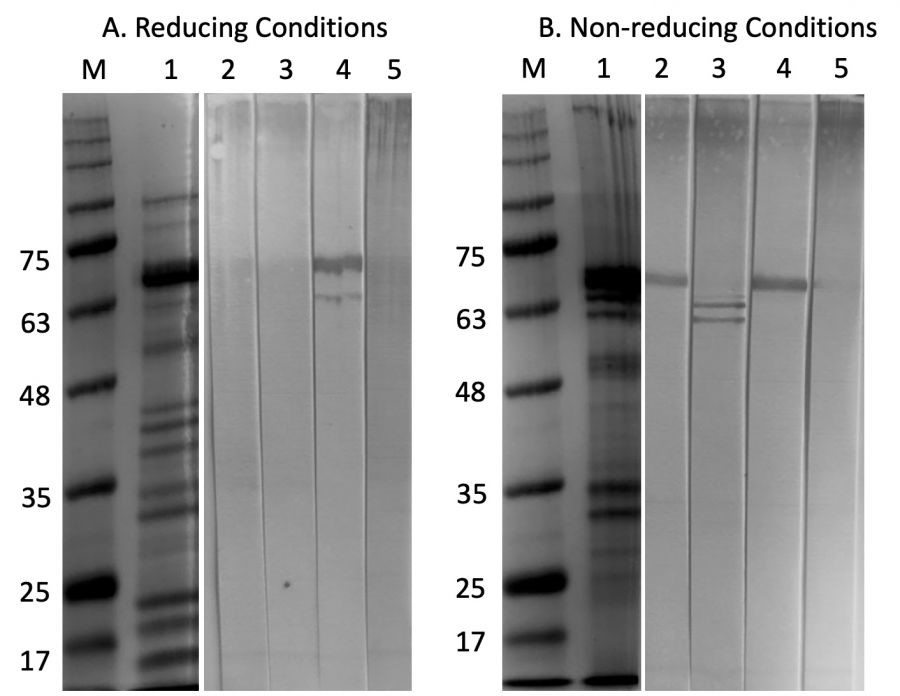
還元条件下(A)および非還元条件下(B)でのCPEのSDS-PAGEおよびイムノブロット解析。
【クーマシー・ブリリアントブルーR-250染色】M:分子マーカー, 1:CPE
【免疫ブロット】2:6E10C12, 3:2G11G7, 4:6B7B10, 5:5E7B12
2)ELISA
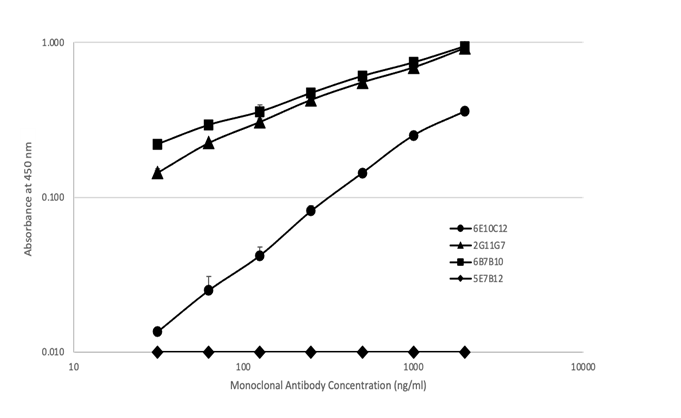
抗マウスIgE抗体でコーティングしたプレートとビオチン化CPEをトレーサーとして使用するサンドイッチELISAで、CPEに対する4種類のマウスIgE mAbを評価しました(図3)。4種類のIgE mAbは、ビオチン化CPEと用量依存的に反応しました。6E10C12、2G11G7、および6B7B10は1~10 ng/mlで同様の反応性を示しましたが、5E7B12は10~100 ng/mlの範囲でCPEに対してより低い反応性を示しました。
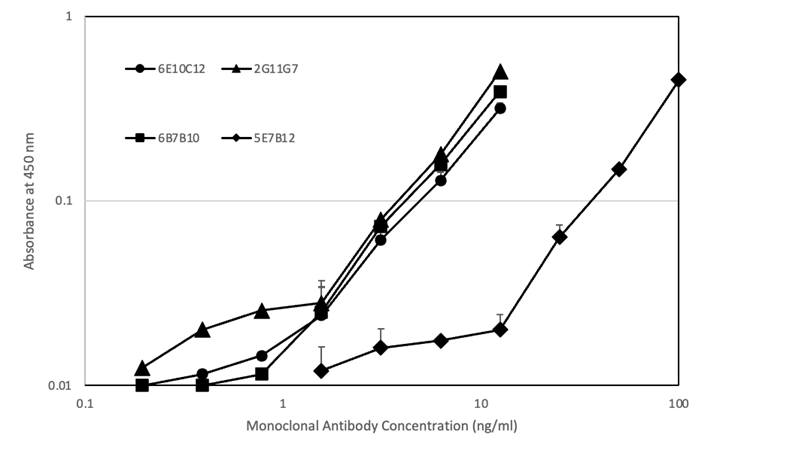
マウス抗CPE IgEモノクローナル抗体(mAb):6E10C12(●)、2G11G7(▲)、6B7B10(■)、5E7B12(◆)をそれぞれ1% BSAを含む0.1Mトリス緩衝食塩水pH7.5で希釈したものを10 μg抗マウスIgEモノクロナル抗体をコーティングしたウェルに加え、室温で2時間インキュベートした。0.05% Tween 20を含む0.05Mリン酸緩衝生理食塩水pH7.4でプレートを洗浄後、2% カゼインを含む0.1Mトリス緩衝生理食塩水pH7.5で希釈したビオチン化CPEを添加し、室温で1時間インキュベートした。その後、洗浄バッファーでプレートを洗浄後、カゼインバッファーで希釈したアビジン-ペルオキシダーゼを加え、室温で30分間インキュベートした。プレートを洗浄後、抗体の結合をTMBで25分間室温で可視化し、停止し、450 nm / 630 nmのOD値を読み取った。
次にCPEをコートしたプレートと抗マウスIgE検出抗体を使用した間接ELISAによって、4つのマウスIgE mAbを評価しました(図4)。間接ELISAでは、2G11G7および6B7B10は、プレート上にコーティングされたCPEに対して、31.5~2000 ng/mlの抗体濃度で、用量依存的な高い反応性を示しました。同じ抗体濃度において、6E10C12はより低い反応性を示し、5E7B12はプレート上にコートされたCPEと反応しませんでした。
マウス抗CPE IgEモノクローナル抗体(mAbs):6E10C12(●)、2G11G7(▲)、6B7B10(■)、5E7B12(◆)をそれぞれ1% BSAを含む0.1Mトリス緩衝食塩水pH7.5で希釈し、1 μgのCPEでコーティングしたウェルに加え、室温で2時間インキュベートした。0.05% Tween20を含む0.05Mリン酸緩衝生理食塩水pH7.4でプレートを洗浄後、2% カゼインを含む0.1Mトリス緩衝生理食塩水pH7.5で希釈したビオチン化抗マウスIgE抗体を添加し、1時間室温でインキュベーションした。その後、洗浄バッファーでプレートを洗浄後、カゼインバッファーで希釈したアビジン-ペルオキシダーゼを加え、室温で30分間インキュベートした。プレートを洗浄後、TMBで25分間室温で反応し、抗体の結合を可視化し、反応停止後、450 nm / 630 nmの吸光度を測定した。
肥満細胞活性化試験
4種類のIgE mAbの生物学的な活性は、RBL-2H3細胞を使用した脱顆粒測定におけるβ-ヘキソサミニダーゼの放出により評価しました。RBL-2H3細胞をマウスIgE mAbの段階希釈(0.128~2000 ng/ml)で感作後、2 μg/mlのCPEで負荷試験を行いました(図5)。釣鐘型の容量曲線で、典型的なプロゾーン効果が観察されました。2G11G7および6E10C12mAbでは、高濃度領域 (400~2000 ng/ml)で比較的低レベルのβ-ヘキソサミニダーゼが放出され、抗体濃度3.2~80 ng/mlにおいて脱顆粒は最大約20%~60%のレベルまで増加しました。0.128 ng/mlの濃度では、β-ヘキソサミニダーゼの放出はバックグラウンドレベルまで減少しました。6B7B10ではβ-ヘキソサミニダーゼ放出レベルは低く、抗体濃度3.2~80 ng/mlにおいて最大15%の脱顆粒を示しました。興味深いことに、5E7B12はRBL-2H3細胞の脱顆粒を引き起こしませんでした。プロゾーン効果は、高濃度のアレルゲンがIgE受容体に結合したIgEと個々に反応し、結果的にアレルゲンによるIgE mAbの架橋を防ぐことに起因します。
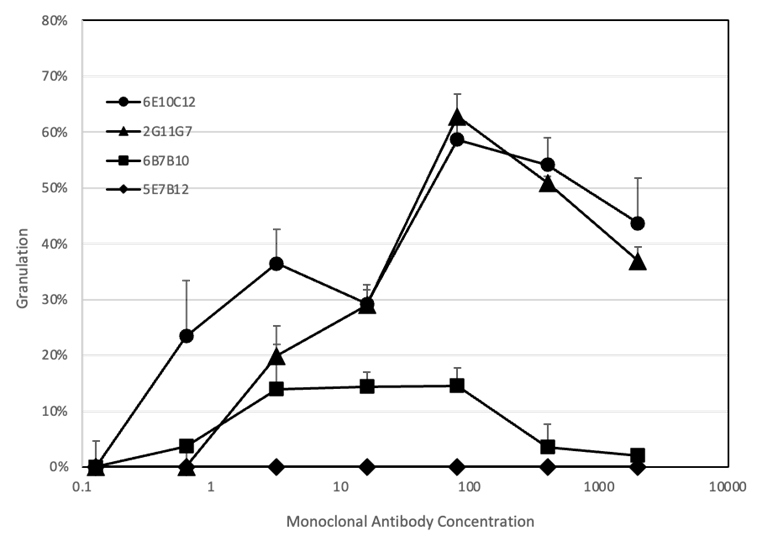
RBL-2H3細胞を15% FBSを含む1 mLのDMEM 、96wellプレート(10^6 細胞/well)37℃で3時間培養し、抗CPE IgEモノクローナル抗体:6E10C12(●)、2G11G7(▲)、6B7B10(■)、5E7B12(◆)で37℃、16時間処理した。PBSで2回細胞を洗浄した後、Tyrode's buffer中の2 μg/mlのCPEを200 μl/wellで添加し、37℃で1時間インキュベートした。その後、各ウェルのサンプル100 μlを96ウェルプレートに移し、β-ヘキソサミニダーゼ活性を測定した。RBL-2H3細胞の脱顆粒は、1% Triton-X Tyrode's bufferを投与した細胞を100%として比率で表した。
次に、RBL-2H3細胞の活性化対するCPE濃度の用量依存性を、3種類のIgE mAbで評価しました(図6)。β-ヘキソサミニダーゼの用量依存的放出は2G11G7および6E10C12(抗体濃度100 ng/ml)ではCPE濃度0.5~8 μg/mlの間で、6B7B10(抗体濃度:20 μg/ml)ではCPE濃度2~8 µg/mlの間で確認されました。データは掲載していませんが、これらのアッセイでは、高濃度の CPE による感作だけでもβ-ヘキソサミニダーゼの放出が確認できました。CPE単独によるRBL-2H3細胞の活性化は、CPEに内在するLPSの存在によって引き起こされている可能性があります。RBL-2H3細胞は細胞表面にTLR2およびTLR4受容体を発現しており、洗浄後もウェルに残っているCD14や MyD88などの血清成分がCPE中のLPSと作用して TLR4受容体を活性化し、結果としてわずかな細胞活性化が起こると考えることができます(15,16)。
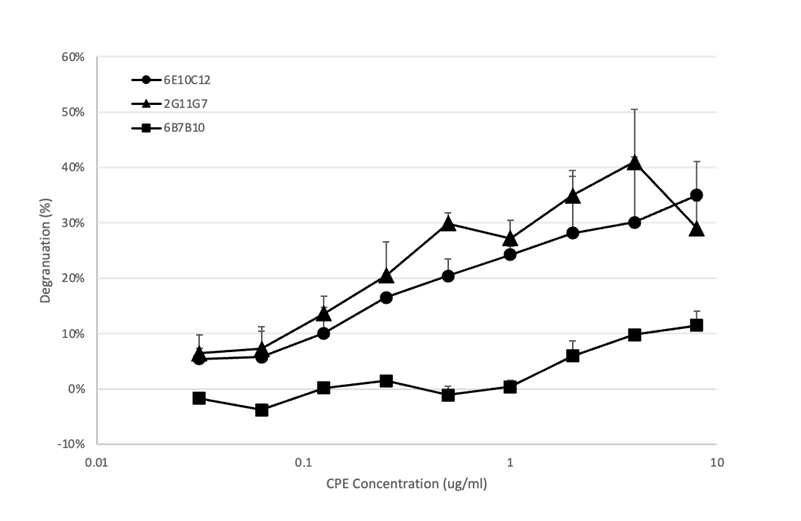
RBL-2H3細胞を15% FBSを含む1 mLのDMEM 、96wellプレート(10^6 細胞/well)37℃で3時間培養し、抗CPE IgEモノクローナル抗体:6E10C12(●)、2G11G7(▲)、6B7B10(■)、5E7B12(◆)で37℃、16時間処理した。PBSで2回細胞を洗浄した後、Tyrode's buffer中の各濃度のCPEを200 μl/wellで添加し、37℃で1時間インキュベートした。その後、各ウェルのサンプル100 μlを96ウェルプレートに移し、β-ヘキソサミニダーゼ活性を測定した。RBL-2H3細胞の脱顆粒は、1% Triton-X Tyrode's bufferを投与した細胞を100%として比率で表した。
動物実験
1)フットパットI型アレルギー反応試験
3つのIgE mAbのマウスI型アレルギー誘導能を評価しました(図7)。2G11G7および6B7B10は、フットパットへのCPE注射の1~2時間後に足の腫れがピークとなり、それぞれ1.10±0.42 mmおよび1.25±0.78 mmに達し、6時間後に元のレベルまで消失しました。ただし、6E10B10は足の腫れを誘導できませんでした。以前の実験で使用されたRBL-2H3細胞は、ラットの好塩基性白血病細胞から樹立されました。マウスIgE抗体はラット細胞のIgE受容体を活性化するものの、6E10B10はマウスのマスト細胞のIgE受容体に対する結合親和性が低い可能性があります。したがって、6E10B10はin vivoでI型アレルギーを誘導できない可能性があります(17)。この抗体の細胞ベースアッセイとマウスにおける研究の矛盾に関しては、さらなる分析が必要です。
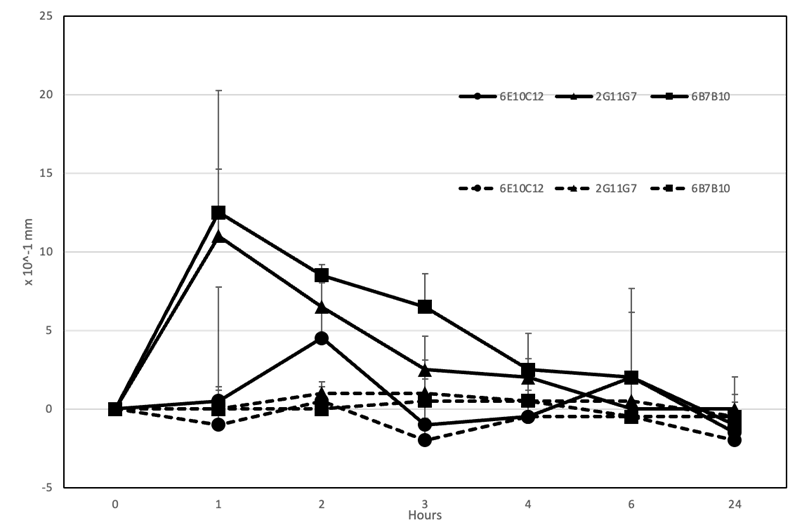
モノクローナル抗体1 mgを静脈内投与した後のマウスの足部の厚さを示した。抗CPE IgEモノクローナル抗体:6E10C12(●)、2G11G7(▲)、6B7B10(■)を投与した後、50μgのCPE(実線)またはPBS(破線)を足蹠部に皮内注射した。
2G11G7によって誘導される足の腫れの用量依存性を評価しました(図8)。100 µg、300 µg、および1000 µg投与時のI型アレルギーの足の腫れは、それぞれ1.30±0.43 mm、1.60±0.13 mm、および2.07±0.10 mmでした。足の腫れはすべての用量で6時間持続し、CPEを足蹠に注射した後8時間以内に元のレベルに減少しました。
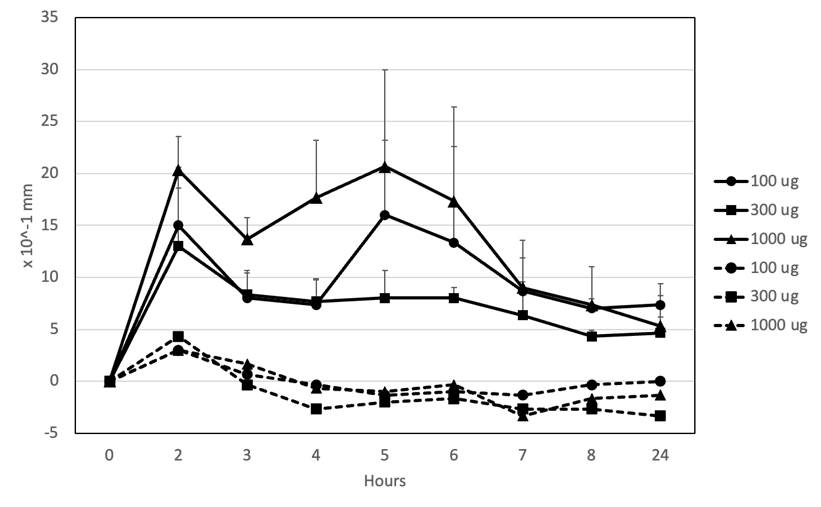
抗CPE IgEモノクローナル抗体2G11G7:100 μg(●)、300 μg(■)、1000 μg(▲)の各用量を静脈内投与した後、100 μgのCPE(実線)またはPBS(破線)を足部に皮内投与した後のマウスの足部の厚みを示す図。
2)マウスアナフィラキシー試験
CPE感作試験では、1 mgの2G11G7または6B7B10を投与されたマウスにおいて、体温がそれぞれ0.9±0.7℃および0.7±0.7℃低下しました。一方、6E10C12は体温低下を誘導できませんでした。体温は90分後に回復しました(図9)。
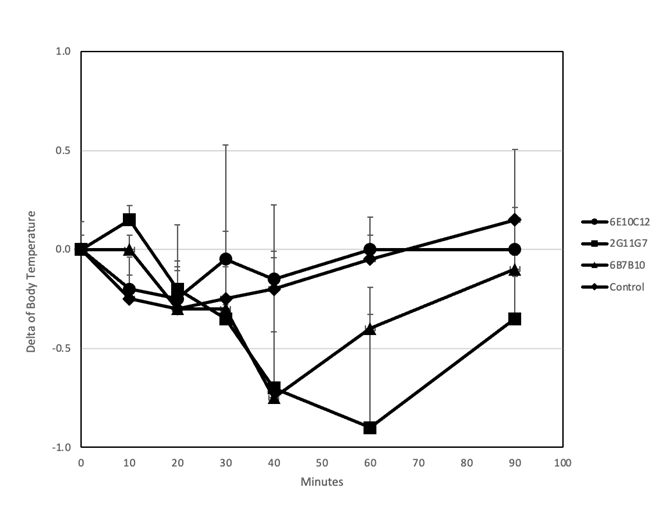
6E10C12(●)、2G11G7(▲)、6B7B10(■)が投与されたマウス。アナフィラキシーを評価するために、抗CPE IgE mAbが投与されたマウスに1 mgのCPEを静脈内投与した後、体温を注射後10分から90分まで直腸で測定した。マウスは免疫の24時間前に、それぞれ1 mgの抗CPE IgE mAbsを静脈内投与で前処理した。
異なる容量の抗CPE IgEモノクローナル抗体2G11G7を静脈注射した後、1 mgのCPEで静脈内投与した結果、10 分後にマウスの体温は1.2~2.7℃低下しました。1000 μg、100 μg、および30 μgの体温低下は、それぞれ2.7±1.7℃、2.1±1.5℃、および1.3±1.6℃でした。体温は30分以内に元のレベルまで回復しました(図10)。
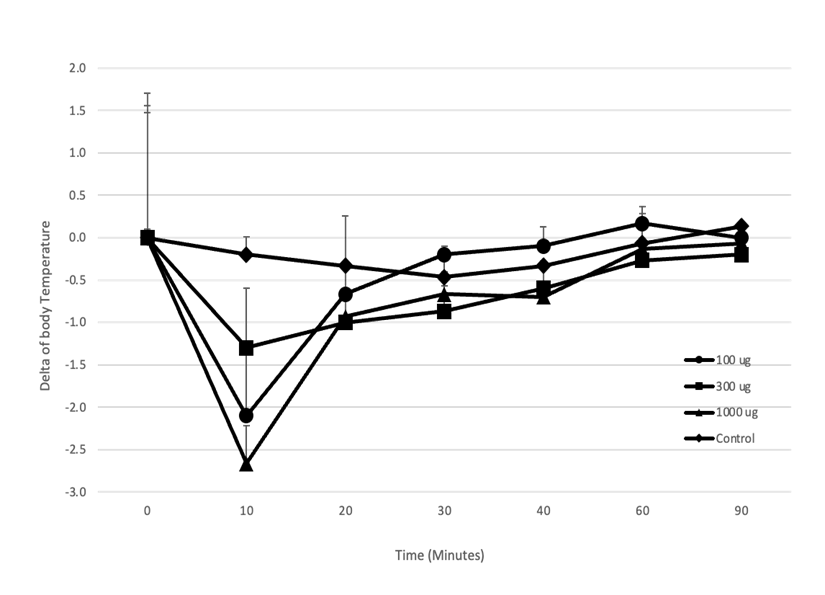
抗 CPE IgEモノクローナル抗体 2G11G7:100 μg(●)、300 μg(■)、1000 μg(▲)の各用量を投与したマウス。アナフィラキシーを評価するために、抗CPE IgE mAbが投与されたマウスに1 mgのCPEを静脈内投与した後、体温を注射後10分から90分まで直腸で測定した。マウスは免疫の24時間前に、それぞれ1 mgの抗CPE IgE mAbsを静脈内投与で前処理した。
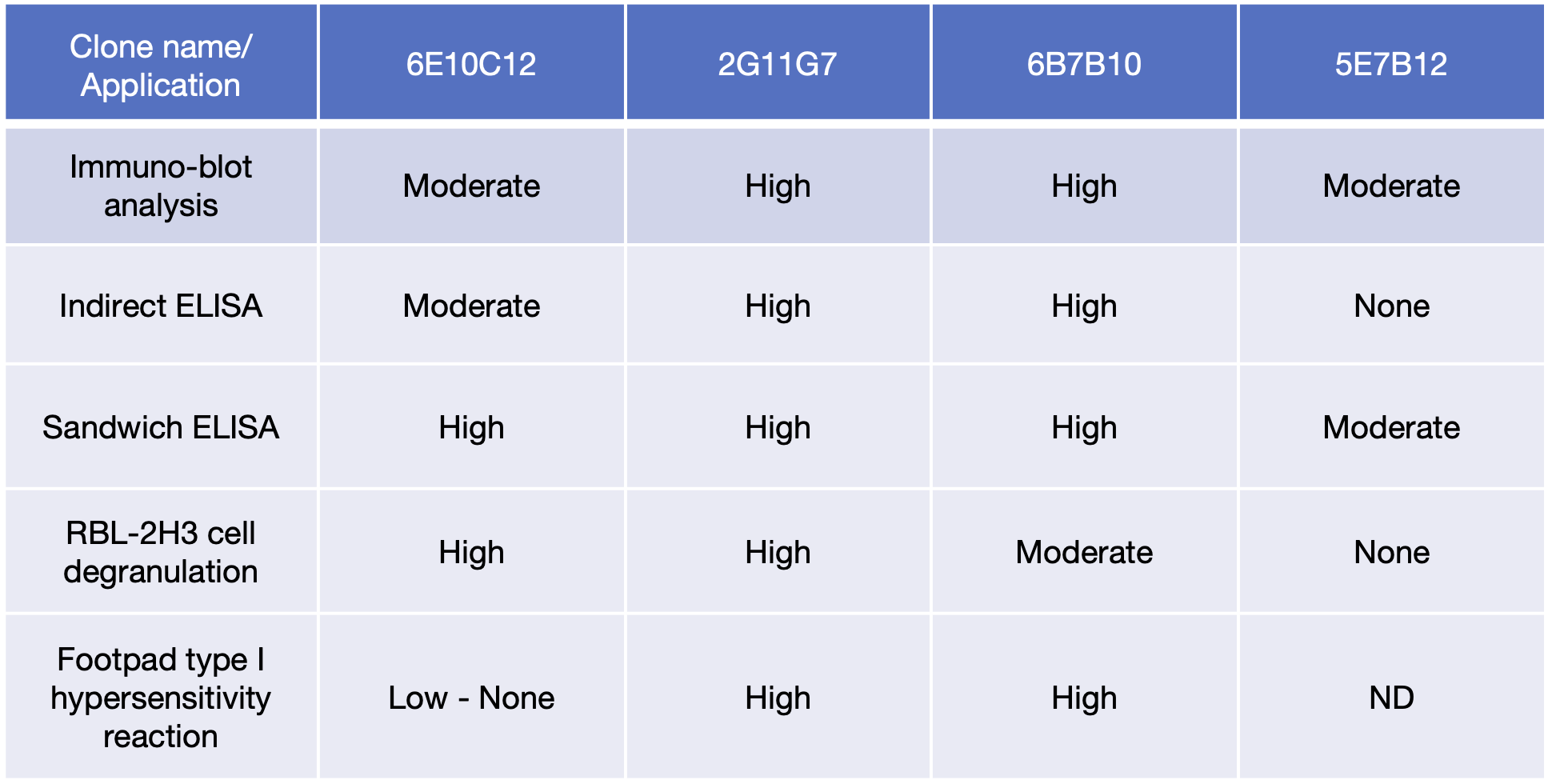
4つのIgEモノクローナル抗体の生物活性のまとめ
参考文献
- A. W. Burks, A. Wesley Burks, Peanut allergy. The Lancet. 371 (2008), pp. 1538–1546.
- Y. Zhuang, S. C. Dreskin, Redefining the major peanut allergens. Immunol. Res. 55, 125–134 (2013).
- J. J. Dolence, Induction of Peanut Allergy Through Inhalation of Peanut in Mice. Methods Mol. Biol. 2223, 19–35 (2021).
- M. K. Selgrade, C. C. Bowman, G. S. Ladics, L. Privalle, S. A. Laessig, Safety assessment of biotechnology products for potential risk of food allergy: implications of new research. Toxicol. Sci. 110, 31–39 (2009).
- X. M. Li, D. Serebrisky, S. Y. Lee, C. K. Huang, L. Bardina, B. H. Schofield, J. S. Stanley, A. W. Burks, G. A. Bannon, H. A. Sampson, A murine model of peanut anaphylaxis: T- and B-cell responses to a major peanut allergen mimic human responses. J. Allergy Clin. Immunol. 106, 150–158 (2000).
- M.-J. Bae, H. S. Shin, E.-K. Kim, J. Kim, D.-H. Shon, Oral administration of chitin and chitosan prevents peanut-induced anaphylaxis in a murine food allergy model. Int. J. Biol. Macromol. 61, 164–168 (2013).
- M. Kulis, X. Chen, J. Lew, Q. Wang, O. P. Patel, Y. Zhuang, K. S. Murray, M. W. Duncan, H. S. Porterfield, A. W Burks, S. C. Dreskin, The 2S albumin allergens of Arachis hypogaea, Ara h 2 and Ara h 6, are the major elicitors of anaphylaxis and can effectively desensitize peanut-allergic mice. Clin. Exp. Allergy. 42, 326–336 (2012).
- L. M. Chang, Y. Song, X.-M. Li, H. A. Sampson, M. Masilamani, Dietary Elimination of Soybean Components Enhances Allergic Immune Response to Peanuts in BALB/c Mice. Int. Arch. Allergy Immunol. 166, 304–310 (2015).
- C. Zhou, T. Ludmila, N. Sun, C. Wang, Q. Pu, K. Huang, H. Che, BALB/c mice can be used to evaluate allergenicity of different food protein extracts. Food Agric. Immunol. 27, 589–603 (2016).
- X. M. Li, D. Serebrisky, S. Y. Lee, C. K. Huang, L. Bardina, B. H. Schofield, J. S. Stanley, A. W. Burks, G. A. Bannon, H. A. Sampson, A murine model of peanut anaphylaxis: T- and B-cell responses to a major peanut allergen mimic human responses. J. Allergy Clin. Immunol. 106, 150–158 (2000).
- C. Tontini, S. Bulfone-Paus, Novel Approaches in the Inhibition of IgE-Induced Mast Cell Reactivity in Food Allergy. Front. Immunol. 12, 613461 (2021).
- H. C. Oettgen, O. T. Burton, IgE receptor signaling in food allergy pathogenesis. Curr. Opin. Immunol. 36, 109–114 (2015).
- O. T. Burton, M. Noval Rivas, J. S. Zhou, S. L. Logsdon, A. R. Darling, K. J. Koleoglou, A. Roers, H. Houshyar, M. A. Crackower, T. A. Chatila, H. C. Oettgen, Immunoglobulin E signal inhibition during allergen ingestion leads to reversal of established food allergy and induction of regulatory T cells. Immunity. 41, 141–151 (2014).
- J. Huang, C. Liu, Y. Wang, C. Wang, M. Xie, Y. Qian, L. Fu, Application of in vitro and in vivo models in the study of food allergy. Food Science and Human Wellness. 7, 235–243 (2018).
- E. Passante, C. Ehrhardt, H. Sheridan, N. Frankish, RBL-2H3 cells are an imprecise model for mast cell mediator release. Inflamm. Res. 58, 611–618 (2009).
- E. Passante, C. Ehrhardt, H. Sheridan, N. Frankish, Toll-like receptors and RBL-2H3 mast cells. Inflamm. Res. 58 Suppl 1, 11–12 (2009).
- A. R. Sterk, T. Ishizaka, Binding properties of IgE receptors on normal mouse mast cells. J. Immunol. 128, 838–843 (1982).

